
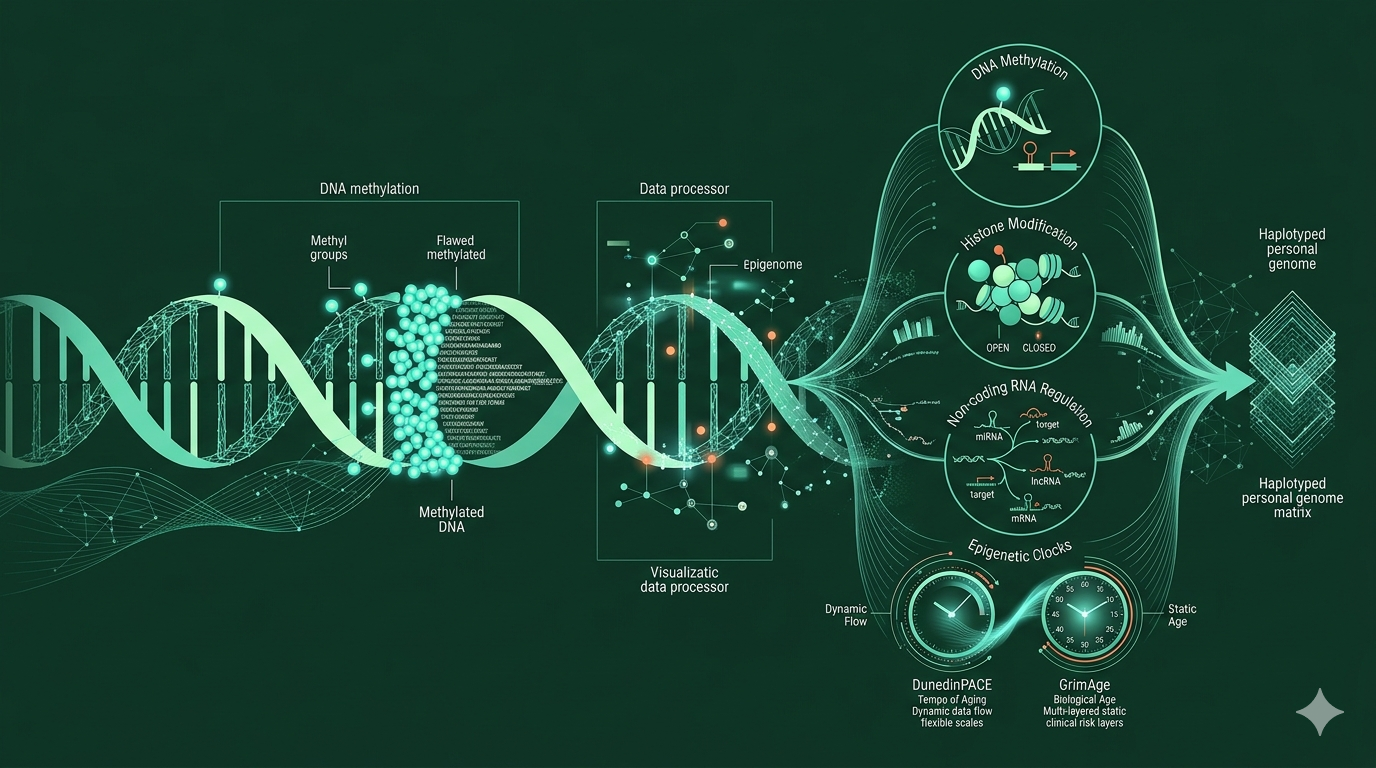
Epigenetika je soubor biologických mechanismů, které určují, jak jsou geny v buňkách zapínány a vypínány, aniž by se měnila samotná sekvence DNA. Mění tedy „čtení“ genetické informace, nikoli její text. Epigenomika je obor, který tyto epigenetické změny studuje v měřítku celého genomu. Zatímco genomika odpovídá na otázku „jaké geny máme", epigenomika se ptá „jak a kdy jsou tyto geny zapínány nebo vypínány" a jak na ně působí životní návyky a podmínky, ve kterých žijeme.
Co je epigentika a proč ji studujeme
DNA v každé buňce lidského těla je prakticky stejná. Přesto se jaterní buňka chová jinak než neuron nebo imunitní buňka. Odpovědí na tento zdánlivý paradox je epigenetika.
Nejlépe prozkoumaným mechanismem je metylace DNA (DNA methylation) — chemická modifikace, při níž se k základním stavebním kamenům DNA (bazím) váže metylová skupina. Tato změna funguje jako záložka v knize genomu: nezmění samotný text, ale signalizuje, zda má být daný úsek čten nebo přeskočen. Dalšími klíčovými mechanismy epigenetické regulace jsou modifikace histonů (proteinů, kolem nichž je DNA namotána v jádře bunky), přístupnost chromatinu (jak moc je DNA v tomto místě „otevřená“ a dostupná pro vazbu proteinů koordinujících přepis genů) a nekódující RNA molekuly jako miRNA (microRNA) nebo lncRNA (long non-coding RNA).
Dohromady tyto prvky tvoří epigenom - soubor všech epigenetických „poznámek“ v buňce, který se v průběhu života mění. Epigenomika se plně rozvinula s příchodem vysokopropustných sekvenačních technologií a čipových platforem. Dnes víme, že epigenomické vzorce se liší mezi populacemi, tkáněmi, mění se s věkem, reagují na vlivy okolního prostředí a životní styl a souvisejí s rizikem chronických onemocnění. Obor se tak postupně posunul od kuriozity laboratoří molekulární biologie k jedné z hlavních os výzkumu preventivní medicíny a biologického stárnutí.
Epigenetické hodiny: biologický věk, který lze měřit
Jedním ze zásadnějších objevů poslední dekády byl vývoj tzv. epigenetických hodin (epigenetic clocks). Jde o matematické modely, které na základě metylačních vzorců na konkrétních místech genomu odhadují biologický věk člověka, tedy stav jeho tkání a orgánů. Lidé stejného kalendářního věku mohou biologicky stárnout různě rychle, a právě to se epigenetické hodiny snaží zachytit.
Průkopnické epigenetické hodiny sestavil Steve Horvath v roce 2013. Z metylačních dat z různých typů tkání navrhl model, který odhaduje věk s vysokou přesností i mezi biologicky odlišnými tkáněmi (Horvath, 2013). Hannum a kolegové ve stejném roce publikovali první epigenetické hodiny konstruované specificky z metylačního profilu krve, které umožňovaly kvantifikovat individuální rychlost stárnutí a ukázaly, že tuto rychlost ovlivňují genetické faktory, pohlaví a pravděpodobně také různé environmentální faktory (Hannum et al., 2013).
Klíčovým krokem vpřed byl vývoj modelu GrimAge (2019): hodiny navržené tak, aby co nejlépe předpovídaly budoucí zdravotní stav a riziko úmrtí. GrimAge kombinuje metylační markery s biologickými signály, jako jsou odhadované hladiny některých proteinů a expozice kouření, a předpovídá délku a kvalitu života i riziko úmrtí z jakékoli příčíny lépe než samotný kalendářní věk. (Lu et al. 2019, McCrory et al., 2021). Přímé srovnání více hodin potvrdilo, že GrimAge je v predikci klinicky významných stavů a celkové úmrtnosti nejúspěšnější (McCrory et al., 2021).
Nejnovějším přírůstkem je DunedinPACE (2022), který namísto odhadu věku měří tempo stárnutí — tedy jak rychle jedinec v daném časovém okamžiku biologicky stárne (Belsky et al., 2022). Pro klinické aplikace je toto rozlišení zásadní, protože zatímco GrimAge říká „kde jsi biologicky dnes“, DunedinPACE říká „jak rychle tam míříš“. Pro hodnocení efektu intervencí (pohybová aktivita, změna diety, odvykání kouření) je dynamické měření tempa potenciálně cennější než statický odhad věku. Studie Belskyho a kolegů ukázala, že DunedinPACE dává stabilní výsledky při opakovaném měření téhož člověka, silně koreluje se zdravotními obtížemi, omezením funkčních schopností a rizikem úmrtí a přidává prediktivní hodnotu i nad rámec GrimAge.
Pro lékaře a výzkumníky mohou být tyto nástroje atraktivní, protože umožňují stratifikovat riziko nemocí nad rámec toho, co říká rodný list. Biologický věk odvozený z epigenetických hodin se jeví jako slibný kandidát na biomarker, který by mohl v budoucnu doplnit stávající klinické ukazatele, ale zatím není jasné, v jaké podobě a v jakých situacích by měl být rutinně používán.
Co epigenom ovlivňuje: kde jsou důkazy nejsilnější
Z velkých epigenomických studií dnes vyplývá, že některé životní návyky a okolní vlivy zanechávají v epigenomu výraznou a opakovaně potvrzenou stopu. Nejsilnejší asociace mezi životním stylem a metylací DNA se ukazují u následujících faktorů:
Kouření je nejlépe zdokumentovaným příkladem modifikovatelného vnějšího faktoru, který zanechává výraznou stopu v epigenomu. Rozsáhlá mezinárodní studie identifikovala více než 2 500 míst v 1 405 genech, jejichž metylace se u kuřáků statisticky významně liší od nekuřáků, přičemž nejsilnější a nejkonzistentnější signál byl nalezen v genu AHRR. AHRR se podílí na detoxifikaci karcinogenů a jeho hypometylace je dnes považována za jeden z nejspolehlivějších biologických markerů kouření (Joehanes et al., 2016). Důležité z hlediska prevence je, že část těchto změn je do značné míry reverzibilní: po zanechání kouření se metylace postupně vrací k hodnotám nekuřáků, i když některé stopy mohou přetrvávat ještě roky jako „biologická paměť“ dřívějšího chování (Gao et al., 2015).
Obezita a s ní spojené poruchy metabolismu patří po kouření k lépe prozkoumaným oblastem epigenomického výzkumu. Wahl a kolegové ve své studii z roku 2017 propojili index tělesné hmotnosti (Body Mass Index, BMI) s metylačními změnami v genech ovlivňujících metabolismus tuků, inzulinovou signalizaci a zánět. Genetické analýzy ukázaly, že tyto změny většinou vznikají jako důsledek vyšší tělesné hmotnosti a s ní souvisejících metabolických poruch, nikoli jako jejich primární příčina. Výjimkou bylo jedno konkrétní místo genomu (u genu NFATC2IP/SH2B1), u něhož bazální úroveň metylace předpovídala budoucí nárůst hmotnosti, což naznačuje možnou příčinnou roli epigenetické regulace alespoň v části případů. Autoři zároveň ukazují, že specifický metylační profil v periferní krvi výrazně předpovídá vznik diabetu 2. typu, a může tak pomoci odlišit metabolicky nepříznivou formu obezity (Wahl et al., 2017). Jiné studie ukazují, že redukce tělesné hmotnosti, například po bariatrické operaci, tj. zmenšení žaludku nebo zkrácení trávicího traktu, byla spojena s částečnou úpravou metylačních vzorců směrem k profilům pozorovaným u lidí se zdravou hmotností, i když efekty nebyly plně reverzibilní (Talukdar et al., 2022; Izquierdo et al., 2019).
Pravidelná fyzická aktivita je spojena s metylačními změnami v kosterním svalu, tukové tkáni a krvi — zejména v drahách pro inzulinovou signalizaci, zánět a mitochondriální funkci (Voisin et al. 2015). Intervenční studie naznačují, že pohyb může biologický věk, jak ho zachycují epigenetické ukazatele, mírně zpomalit, ale přesna “dávka” a velikost efektu se napříč studiemi liší a zůstává předmětem výzkumu.
Podobně je tomu se stravou. Nejsolidnejší důkazy existují pro tzv. metyl‑donorové živiny – folát, vitamin B12, cholin a methionin – které přímo zásobují biochemické reakce metylace DNA (Joubert et al., 2016). Středomořský stravovací vzorec, tedy méně kalorická strava a potraviny bohaté na polyfenoly vykazují slibné asociace s metylací a epigenetickým věkem. Efekty jsou však v populačních studiích obvykle mírné a výrazně závislé na kontextu a sledované populaci (Fiorito et al., 2019). Joubert a kolegové (2016) navíc prokázali, že hladina folátu v mateřské plazmě v těhotenství ovlivňuje metylaci u novorozenců, což spojuje stravovací návyky matky s epigenetickým programováním potomka (Joubert et al., 2016).
Stres a spánek mají v epigenetickém výzkumu zvláštní postavení. Několik kohortových studií ukazuje, že narušený cirkadiánní rytmus a dlouhodobý nedostatek spánku souvisejí s metylačními změnami a rychlejším epigenetickým stárnutím (Lee et al., 2024; Rivero‑Segura et al., 2025). Analýza chronického psychosociálního stresu v populační kohortě popsala asociace mezi dlouhodobým stresem a změnami metylace na vybraných místech v genech souvisejících se stresem (Miller et al., 2026). Většina prací zabývající se těmito faktory je však observačních, používá různé definice stresu i spánkových ukazatelů a různé typy epigenetických hodin, takže výsledky je třeba interpretovat opatrně.
Rané životní podmínky jsou dalším specifickým tématem epigenomického výzkumu. Studie Heijmanse a kolegů publikovaná již v roce 2008 prokázala, že nedostatek výživy u těhotných žen v průběhu „holandské hladové zimy“ v letech 1944–45 zanechal u potomků trvalé metylační změny v genu IGF2, které byly detekovatelné ještě desítky let po narození (Heijmans et al., 2008). Tento nález otevřel výzkum toho, jak podmínky v raném vývoji – od výživy přes stres až po sociální prostředí – formují epigenetické nastavení organismu a ovlivňují zdravotní rizika v dospělosti. Navazující práce ukázaly, že různé formy negativních podmínek v dětsví souvisejí se zvýšenou epigenetickou záteží ve středním a vyšším věku (McCrory et al., 2022; Joshi et al., 2023). Zároveň platí, že jde převážně o observační kohorty, takže přesný rozsah a mechanizmy těchto efektů jsou stále předmětem výzkumu.
Co to znamená v praxi: pro pacienty i lékaře
V praxi zatím u vetšiny klinických oblastí epigenomika nestojí jako samostatný screeningový nebo diagnostický nástroj, ale vývoj jde tímto směrem. Výjimku představuje onkologie, kde analýza epigenetických markerů nádorů již slouží k jejich klasifikaci, diagnostice a sledování léčby (mimo jiné prostřednictvím tekuté biopsie). Nicméně, jde o oblast nádorové medicíny, nikoli preventivní epigenomiky. Mezi dalších několik oblastí, kde je klinický potenciál nejvýraznější patří:
Stratifikace kardiovaskulárního a metabolického rizika. Biologický věk odvozený z metylace by mohl v budoucnu přispět k přesnějšímu posouzení rizika nad rámec klasických parametrů jako cholesterol, krevní tlak nebo glykémie, což naznačují práce, kde epigenetické hodiny předpovídají úmrtnost a klinické fenotypy lépe než samotný kalendářní věk. Zatím však jde spíše o oblast výzkumu než standardní součást stratifikace pacientů.
Monitorování efektu změny životního stylu. Epigenetické hodiny — zejména DunedinPACE — by mohly potenciálně sloužit jako objektivní biomarker toho, zda pacient biologicky odpovídá na změnu životního stylu, např. při odvykání od kouření nebo redukci hmotnosti. Klinické studie v tomto směru probíhají, ale přímé a jednoznačné doporučení pro běžnou praxi zatím chybí.
Prenatální a vývojová medicína. Výsledky Heijmansovy studie a navazujícího výzkumu posilují argumenty pro péči o prenatální výživu, omezení nepříznivých faktorů v těhotenství (kouření, alkohol, znečištění ovzduší) a kvalitní prenatální péči, protože ukazují, že podmínky v těhotenství a raném dětství mohou zanechat dlouhodobé epigenetické stopy s dopadem na zdraví v dospělosti.
Pro pacienta je klíčové sdělení prosté: prostředí a životní styl zanechávají měřitelné biologické stopy. To dává biologický základ smysluplnosti prevence — ale neznamená, že každá epigenetická změna je škodlivá nebo kauzální, ani že lze epigenom libovolně „přepsat" zdravými návyky bez dalších implikací.
Limity a otevřené otázky
Epigenomika je fascinující, ale stále mladý obor s řadou nevyřešených problémů.
Asociace versus kauzalita. Velká část nálezů popisuje korelaci, nikoli prokázanou příčinu. Mnohé metylační změny jsou spíše důsledkem nemoci nebo expozice určitému faktoru prostředí než jejich příčinou. Rozlišit, zda marker mechanisticky přispívá k patogenezi nebo je pouhým biologickým záznamem expozice, vyžaduje metodicky náročné přístupy (Lappalainen et al., 2017).
Složení buněk jako hlavní metodologický problém. Krevní metylace je silně ovlivněna relativním zastoupením jednotlivých typů bílých krvinek, které se mění s věkem, nemocí i jinými faktory. Studie bez korekce na složení buněčné populace mohou generovat falešné asociace. (Jaffe et al., 2014)
Přenositelnost modelů napříč populacemi. Epigenetické hodiny a jiné experimentální modely byly převážně trénovány na evropských kohortách. Jejich výkon v jiných populacích — jiných etnických skupinách, geografických oblastech nebo socioekonomických kontextech — nemusí být stejný. Tento problém se aktivně řeší, ale není dosud vyřešen.
Spolehlivost a reprodukovatelnost. Technické variace mezi laboratořemi a volba metody normalizace výrazně ovlivňují výsledky. Higgins-Chen a kolegové (2022) ukázali, že technické faktory mohou hodnoty epigenetických hodin výrazně posunout a navrhli způsoby korekce pro longitudinální a klinické využití (Higgins-Chen et al., 2022).
Etické otázky. Pokud se biologický věk stane dostupným klinickým testem, vyvstanou otázky o komunikaci výsledků pacientům, o riziku diskriminace při pojišťování nebo zaměstnání a o ochraně epigenetických dat. Tyto otázky nejsou systémově řešeny na regulatorní úrovni.
Závěr
Epigenomika přináší přesvědčivé důkazy o tom, že životní styl a prostředí zanechávají měřitelné biologické stopy v podobě metylačních vzorců, které ovlivňují riziko nemocí a tempo biologického stárnutí. Nejlépe zdokumentovanými modifikovatelnými faktory jsou kouření, obezita, fyzická aktivita a prenatální expozice. Epigenetické hodiny jsou slibným nástrojem pro posouzení biologického věku nad rámec kalendářního věku, ale jejich klinická standardizace naráží na otázky reprodukovatelnosti, populační přenositelnosti a chybějících intervenčních studií s tvrdými klinickými výsledky. Pro lékaře i pacienty z toho plyne jasné poselství: prevence a zdravý životní styl mají biologicky měřitelný efekt — i když epigenom není jednoduše a libovolně přeprogramovatelný.
Zdroje
Belsky, D. W., Caspi, A., Corcoran, D. L., et al. (2022). DunedinPACE, a DNA methylation biomarker of the pace of aging. eLife, 11, e73420. https://doi.org/10.7554/eLife.73420
Fiorito, G., McCrory, C., Robinson, O., et al. (2019). Socioeconomic position, lifestyle habits, and biomarkers of epigenetic aging: a multi-cohort analysis. Aging, 11(7), 2045–2070. https://doi.org/10.18632/aging.101895
Gao, X., Jia, M., Zhang, Y., et al. (2015). DNA methylation changes of whole blood cells in response to active smoking exposure in adults: a systematic review. Clinical Epigenetics, 7, 113. https://doi.org/10.1186/s13148-015-0148-3
Hannum, G., Guinney, J., Zhao, L., et al. (2013). Genome-wide methylation profiles reveal quantitative views of human aging rates. Molecular Cell, 49(2), 359–367. https://doi.org/10.1016/j.molcel.2012.10.016
Heijmans, B. T., Tobi, E. W., Stein, A. D., et al. (2008). Persistent epigenetic differences associated with prenatal exposure to famine in humans. PNAS, 105(44), 17046–17049. https://doi.org/10.1073/pnas.0806560105
Higgins-Chen, A. T., Thrush, K. L., Wang, Y., et al. (2022). A computational solution for bolstering reliability of epigenetic clocks: Implications for clinical trials and longitudinal tracking. Nature Aging, 2, 644–661. https://doi.org/10.1038/s43587-022-00248-2
Horvath, S. (2013). DNA methylation age of human tissues and cell types. Genome Biology, 14(10), R115. https://doi.org/10.1186/gb-2013-14-10-r115
Izquierdo, A. G., & Crujeiras, A. B. (2019). Obesity-Related Epigenetic Changes After Bariatric Surgery. Frontiers in Endocrinology, 10, 232. https://doi.org/10.3389/fendo.2019.00232
Jaffe, A. E., & Irizarry, R. A. (2014). Accounting for cellular heterogeneity is critical in epigenome-wide association studies. Genome Biology, 15(2), R31. https://doi.org/10.1186/gb-2014-15-2-r31
Joehanes, R., Just, A. C., Marioni, R. E., et al. (2016). Epigenetic signatures of cigarette smoking. Circulation: Cardiovascular Genetics, 9(5), 436–447. https://doi.org/10.1161/CIRCGENETICS.116.001506
Joubert, B. R., den Dekker, H. T., Felix, J. F., et al. (2016). Maternal plasma folate impacts differential DNA methylation in an epigenome-wide meta-analysis of newborns. Nature Communications, 7, 10577. https://doi.org/10.1038/ncomms10577
Joshi, D., Gonzalez, A., Lin, D., & Raina, P. (2023). The association between adverse childhood experiences and epigenetic age acceleration in the Canadian longitudinal study on aging (CLSA). Aging Cell, 22(2), e13779. https://doi.org/10.1111/acel.13779
Lappalainen, T., & Greally, J. M. (2017). Associating cellular epigenetic models with human phenotypes. Nature Reviews Genetics, 18(7), 441–451. https://doi.org/10.1038/nrg.2017.32
Lee, H. S., Kim, B., & Park, T. (2024). The association between sleep quality and accelerated epigenetic aging with metabolic syndrome in Korean adults. Clinical Epigenetics, 16, 92. https://doi.org/10.1186/s13148-024-01706-x
Lu, A. T., Quach, A., Wilson, J. G., et al. (2019). DNA methylation GrimAge strongly predicts lifespan and healthspan. Aging, 11(2), 303–327. https://doi.org/10.18632/aging.101684
McCrory, C., Fiorito, G., Hernández, B., et al. (2021). GrimAge outperforms other epigenetic clocks in the prediction of age-related clinical phenotypes and all-cause mortality. The Journals of Gerontology: Series A, 76(5), 741–749. https://doi.org/10.1093/gerona/glaa286
McCrory, C., Fiorito, G., O’Halloran, A. M., Polidoro, S., Vineis, P., & Kenny, R. A. (2022). Early life adversity and age acceleration at mid-life and older ages indexed using the next-generation GrimAge and Pace of Aging epigenetic clocks. Psychoneuroendocrinology, 137, 105643. https://doi.org/10.1016/j.psyneuen.2021.105643
Miller, M., Brown, R. D., Krautkramer, C., et al. (2026). Association between chronic stress and the epigenome: Exploration of psychological and biological stress. PLOS ONE, 21(4), e0346517. https://doi.org/10.1371/journal.pone.0346517
Rivero-Segura, N. A., Cuartas, J. D. R., Garcia-delaTorre, P., Sanchez-Garcia, S., Ramirez-Aldana, R., & Gomez-Verjan, J. C. (2025). Insomnia accelerates the epigenetic clocks in older adults. GeroScience, 47(6), 6777–6788. https://doi.org/10.1007/s11357-025-01608-7
Talukdar, F. R., Escobar Marcillo, D. I., Laskar, R. S., et al. (2022). Bariatric surgery-induced weight loss and associated genome-wide DNA-methylation alterations in obese individuals. Clinical Epigenetics, 14, 176. https://doi.org/10.1186/s13148-022-01401-9
Voisin, S., Eynon, N., Yan, X., & Bishop, D. J. (2015). Exercise training and DNA methylation in humans. Acta Physiologica, 213(1), 39–59. https://doi.org/10.1111/apha.12414
Wahl, S., Drong, A., Lehne, B., et al. (2017). Epigenome-wide association study of body mass index, and the adverse outcomes of adiposity. Nature, 541, 81–86. https://doi.org/10.1038/nature20784





